
Badanie 3. Wielkość strumienia migracji.
Model z badania drugiego rozszerzono o stałą częstotliwość migracji równą 10. W tabeli 5.4 znajdują się wyniki obliczeń. Pierwsza liczba oznacza całkowitą liczbę migrantów, zaś w nawiasie umieszczono procentowy udział migrantów w populacji.
Najlepsze wyniki uzyskał model z rozmiarem migracji wynoszącym 9,6%. Mniejsze rozmiary migracji nie są w stanie znacząco wpłynąć na przebieg zbieżności, a za duże migracje zmieniają jej przebieg w sposób zbyt silny. Skolicki i DeJong określają rozmiar 10% jako "niepotrzebnie duży", ale wskazują, że inne źródła podają ten rozmiar jako wartość wyjściową [36]. Prawdopodobnie dla każdego modelu optymalny rozmiar strumienia jest nieco inny i wymaga dostrojenia. Można jednak przyjąć rozmiar 10% jako wartość domyślną.
W niniejszym badaniu model dynamicznego dostrajania rozmiaru migracji (Hong) przedstawiony w podsekcji 2.2.4 się nie sprawdził. Uzyskał porównywalne z innymi modelami wyniki dla małych rozmiarów problemów, lecz dla dużych okazał się nieskuteczny. Przykładowy przebieg rozmiaru migracji znajduje się na rysunku 5.4.
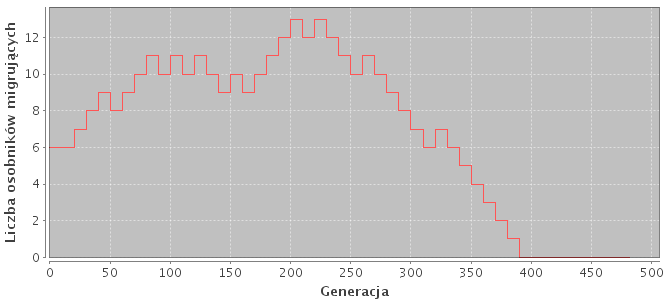
Rysunek 5.4: Dynamiczne dostrajanie rozmiaru migracji. Model zaproponowany przez Hong"a.
Przy ustalaniu rozmiaru migracji należy także wziąć pod uwagę fakt, iż migracja zmniejsza różnorodność populacji. W standardowym modelu migranci są nie przemieszczani, a kopiowani i następnie zastępują osobniki w docelowych podpopulacjach. Następuje zwiększenie liczby genów migrantów kosztem genów osobników wybranych do usunięcia.
Tablica 5.4: Wpływ liczebności strumienia migracji na względną różnicę jakości - D
| Instancja | 32 (1,6%) | 64 (3,2%) | 128 (6,4%) | 192 (9,6%) | 256 (12,8%) | 512 (25,6%) | Hong |
| eil51 | 5.07 | 5.92 | 5.16 | 4.23 | 5.77 | 4.32 | 6.34 |
| berlin52 | 7.21 | 6.89 | 4.51 | 4.95 | 4.56 | 5.24 | 5.52 |
| st70 | 4.30 | 6.76 | 6.22 | 4.06 | 4.59 | 7.91 | 5.13 |
| eil76 | 9.11 | 7.25 | 7.51 | 6.80 | 8.10 | 9.44 | 5.84 |
| pr76 | 4.94 | 4.44 | 5.87 | 3.59 | 6.33 | 5.43 | 4.52 |
| rat99 | 10.57 | 10.70 | 11.45 | 8.84 | 10.01 | 11.25 | 10.75 |
| kroAlOO | 8.89 | 5.39 | 9.09 | 8.90 | 9.29 | 11.36 | 7.17 |
| kroBlOO | 9.17 | 6.78 | 8.40 | 6.01 | 8.46 | 8.68 | 8.42 |
| kroClOO | 8.38 | 10.34 | 9.52 | 8.80 | 6.80 | 8.62 | 8.14 |
| kroDlOO | 7.21 | 7.59 | 8.73 | 7.89 | 6.84 | 7.91 | 9.26 |
| kroElOO | 5.71 | 7.23 | 7.75 | 5.67 | 7.29 | 8.06 | 8.84 |
| rdlOO | 9.53 | 10.76 | 9.13 | 8.78 | 8.96 | 7.66 | 10.98 |
| eillOl | 10.91 | 9.38 | 9.32 | 8.68 | 10.21 | 8.33 | 9.92 |
| linl05 | 8.15 | 5.53 | 7.21 | 8.78 | 9.60 | 5.36 | 9.32 |
| prl07 | 8.09 | 8.16 | 7.88 | 6.96 | 8.82 | 7.92 | 9.1 |
| prl24 | 7.56 | 8.09 | 6.96 | 5.48 | 5.11 | 5.58 | 7.85 |
| bierl27 | 8.20 | 8.59 | 8.17 | 10.52 | 6.90 | 9.19 | 10.04 |
| chi 30 | 11.26 | 9.64 | 8.98 | 7.91 | 8.98 | 8.81 | 9.43 |
| prl36 | 10.41 | 10.17 | 9.27 | 9.94 | 11.14 | 8.38 | 13.31 |
| prl44 | 18.05 | 7.72 | 11.17 | 5.09 | 7.26 | 10.39 | 12.35 |
| chl50 | 19.53 | 13.78 | 12.75 | 12.27 | 14.61 | 12.29 | 13.59 |
| kroA150 | 16.88 | 11.48 | 9.73 | 9.55 | 10.93 | 10.72 | 13.54 |
| kroB150 | 12.46 | 10.33 | 9.36 | 10.46 | 10.39 | 10.17 | 11.3 |
| ul59 | 17.94 | 13.88 | 16.75 | 15.51 | 15.09 | 12.98 | 19.72 |
| S´rednia | 9.98 | 8.62 | 8.79 | 7.9 | 8.59 | 8.58 | 9.6 |
komentarze
Copyright © 2008-2010 EPrace oraz autorzy prac.